


I sommer har NRK gjengitt påstander fra Gjermund Andersen i Naturvernforbundets skogutvalg om at den genetiske diversiteten i foredlet gran er veldig smal og at dette gir opphav til lite robust skog. Vi kan røpe at dette ikke er riktig. Heldigvis.
TEKST OG FOTO: TOR MYKING, NIBIO


I denne artikkelen vil vi kort gjennomgå den forskningen som underbygger dette synet.
Det er bra at det er interesse for genetisk diversitet i gran som plantes. Etablering av ny skog innebærer et stort ansvar
– både av hensyn til biomangfold, til skognæringen og til allmennhetens bruk og interesser. I tillegg har det en etisk dimensjon. Genetisk diversitet er råmateriale for evolusjonen, og dersom den genetiske diversiteten strupes står skogen dårligere rustet til å møte fremtidens klima og miljø. Ingen er tjent med det.
STARTET PÅ 40-TALLET
Skogplanteforedlingen i Norge startet i det små allerede på 1940-tallet. På det tidspunktet var skogvolumet drøyt en tredjedel av dagens nivå (32 % i 1933 og 43 % i 1967), og skogplanteforedling var et av mange tiltak som skulle bidra til å styrke skognæringen gjennom plantemateriale med bedre tilvekst, kvalitet og klimatilpasning. I de første tiårene ble det samlet podekvist fra flere tusen plusstrær, altså skade frie trær med god vekst og gode kvalitetsmessige egenskaper. Podekvistene ble podet på grunnstammer i førstegenerasjons frøplantasjer som etter noen år begynte å produsere frø.
Mange av frøplantasjene ble anlagt tidlig på 1960-tallet, og flere kom til i påfølgende tiår. Andelen foredlet frø brukt i granplantinger har økt jevnt og trutt og er nå over 90 %, som svarer til ca. 35 millioner granplanter per år.
VEISKILLE FØRTE TIL STUDIUM
For rundt 15 år siden begynte arbeidet med å planlegge andregenerasjons frøplantasjer, og skogplanteforedlingen sto derfor ved et veiskille. Hvordan skulle utvalget for neste generasjons frøplantasjer gjøres slik at både genetisk diversitet og genetisk gevinst ble ivaretatt? Dette spørsmålet var utgangs punktet for NFR-prosjektet SustBreed som hadde til hensikt å undersøke genetisk diversitet i plantematerialet fra førstegenerasjons frøplantasjer.
Vi valgte ut to frøplantasjer med forskjellig antall kloner, Kaupanger frøplantasje i Sogn med 25 kloner og Kilen frø plantasje i Telemark med 60 kloner, og studerte genetisk diversitet i frø fra både gode og dårlige frøår.
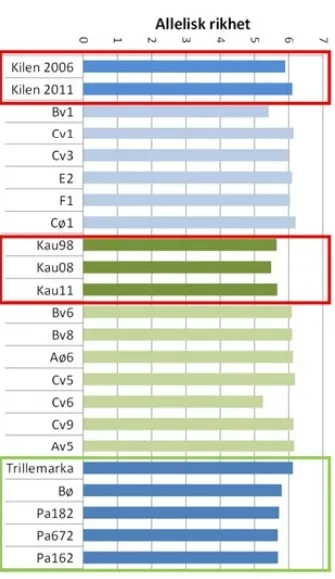
ALLERISK RIKHET
Allelisk rikhet et meget følsomt mål på genetisk variasjon, og resultatene viser at genetisk diversitet i frø fra de to frø plantasjene er innenfor det naturlige variasjonsområdet for gran. For eksempel finnes det bestandsfrø som både har lavere (sankeområdene Bv1/ Cv6) og høyere genetisk diversitet (sankeområdene Cø1/ Cv5) enn frøet fra de to frøplantasjene. Samtidig ser det ut til å være en effekt av antall kloner i frø plantasjene. Kaupanger med sine 25 kloner har konsistent litt lavere genetisk diversitet enn Kilen med 60 kloner. Hoved inntrykket er imidlertid at frøplantasjefrø har en genetisk diversitet på nivå med skog som har opprinnelse i naturlig foryngelse. En nylig svensk studie på gran og furu fant svakt lavere genetisk diversitet på frø fra frøplantasjer enn fra naturpopulasjoner (Heuchel et al. 2022).
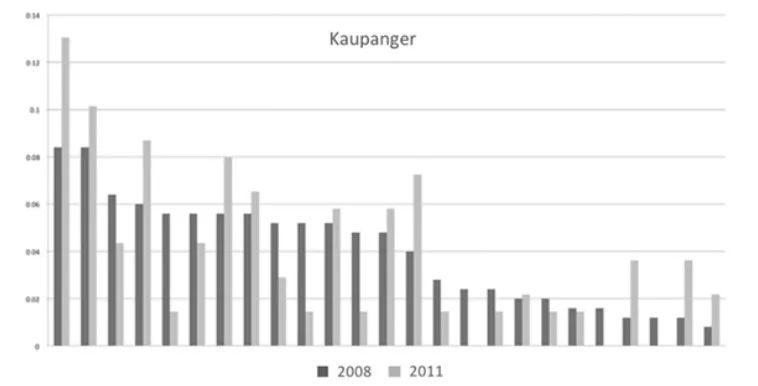
Som vi ser i Figur 2 er det stor forskjell i pollenbidrag fra ulike fedre i Kaupanger frøplantasje. Noen bidrar mye, andre lite, og forskjellen mellom kloner virker ganske konsistent over tid. Likeledes er det stor forskjell i frø produksjonen hos ulike mødre. Begge deler bidrar til å redusere det vi kaller den effektive populasjonsstørrelsen (Ne) som uttrykker hvor mange individer som faktisk bidrar i reproduksjonen. Slektskap kan også bidra til redusert effektiv populasjonsstørrelse. Jo sterkere slektskap i en populasjon, desto lavere effektiv populasjonsstørrelse.
50 UBESLEKTEDE INDIVIDER ER NOK
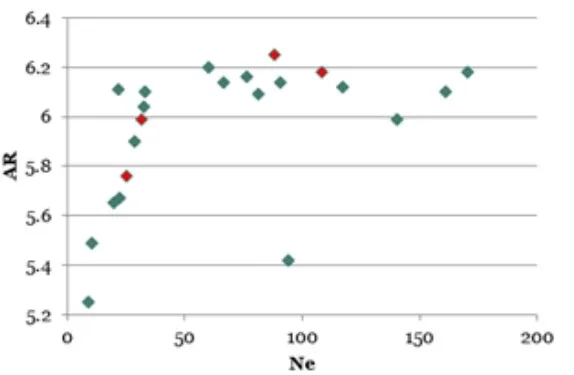
I en ideell populasjon vil 50 ubeslektede individer dekke det aller meste av den genetiske variasjonen. I samsvar med denne teoretiske forventningen ser vi i vår studie at den genetiske diversiteten flater ut ved en effektiv populasjons størrelse på 35–50 (Figur 3), dette ved en observert fremmed pollinering på drøyt 20 %. Fremmedpollineringen var sterkest i dårlige frøår, og den bidrar både til å redusere den genetiske gevinsten og til å øke den genetiske diversiteten (og Ne). Uten fremmedpollinering vil utflatingen skjedd ved noe et høyere klontall. Vi kan imidlertid regne med at alle frøplantasjer har en viss fremmedpollinering, slik at utflatingen av effektiv populasjonsstørrelse på 35–50 kloner kan antas å gjelde generelt.
GRENSER FOR KLONTALL
Som nevnt er det vanlig at den effektive populasjonsstørr elsen ligger godt under klontallet i en frøplantasje, eller i hvilken som helst populasjon. I vår undersøkelse (Sønstebø mfl. 2018) var den effektive populasjonsstørrelsen på knapt 70 % av det aktuelle klontallet i frøplantasjene. Dersom den effektive populasjonsstørrelsen skal ligge på 35–50 (Figur 3) må klontallet derfor være på 45–75, som altså er mer likt klontallet i Kilen frøplantasje enn i Kaupanger frøplantasje. Skogfrøverket har satt grenser for klontall i andregenerasjons frøplantasjer i samsvar med funnene fra disse undersøkelsene.
GENOMISK VERKTØY
Resultatene gjengitt i Figur 1-3 er fra såkalte mikrosatellitter (SSR), lengdevariasjon i korte DNA-sekvenser på i alt 11 steder i genomet. Dette er selvsagt en begrensning
– for hva med variasjonen andre steder i genomet? Siden den under søkelsen (SustBreed) ble foretatt har genomiske verktøy blitt vanligere. Med mange tusen godt fordelte markører gir genomiske analyser et langt fyldigere bilde av den genetiske variasjonen. Genomiske data fra en studie som foreløpig ikke er publisert viser at første seleksjonstrinn (plusstrær) ikke reduserte diversiteten utover det vi observerte i naturlige populasjoner i Norge. Lavest genetisk diversitet hadde naturpopulasjonene i utkanten av utbredelsen, for eksempel på Vestlandet. For andre europeiske treslag er bildet mye det samme, nesten all genetisk diversitet i naturpopulasjoner er ivaretatt etter første seleksjonstrinn (Olsson mfl. 2023).
«FOREDLET SKOG» ER LIKE ROBUST
Ut fra de studiene vi har gjort så langt og utvalgsarbeidet som ellers gjøres i Norge er det heller ikke holdepunkter for å anta at skog med opprinnelse i foredlet frø er mindre robust og får mer klimaskader enn skog med bakgrunn i frø fra naturskog eller andre bestander. Tvert imot, kan vi vel si. I tillegg til at genetisk diversitet er opprettholdt er det etablert et godt system for å fremme at ulike plantematerialer blir brukt på riktig sted, slik at det blir et godt samsvar mellom plantenes vekstrytme og klimaet på voksestedet. Her passer det å nevne Proveniensvelgeren (www.skogfroverket.no/ veiledning/) som veileder frøkjøpere i alternative plantematerialer for spesifikke bruksområder.
TILPASNING TIL KLIMAENDRINGER
En bekymring vi deler med både fagfolk og lekfolk i inn- og utland er at klimaendringene vil foregå raskere enn trærne kan tilpasse seg, blant annet på grunn av lang generasjons tid. Dette kan innebære ulike typer klimaskader, tap av vitalitet og i en del tilfeller at trærne dør. I en slik situasjon kan målrettet bruk av karakteriserte plantematerialer være særlig viktig, noe som i praksis vil være å bruke plante materiale som er tilpasset et varmere klima med lenger vekstsesong enn lokalt plantemateriale. I en slik situasjon er det avgjørende med tilstrekkelig tilgang på passende plante materiale og kunnskap om hvor de bør brukes.
NYTT SELEKSJONSTRINN
Som vist over for gran innebærer skogplanteforedling en mindre innsnevring av den genetiske diversiteten sammenlignet med frø fra bestandsfrø. Ved etablering av første generasjons frøplantasjer skjedde dette som nevnt ved pluss treutvalget. Disse frøplantasjene har senere blitt tynnet, assistert av resultater fra avkomforsøk, slik at de inneholder relativt mer av de beste klonene. Etablering av andregenerasjons frøplantasjer som foregår nå er et nytt seleksjonstrinn. I tillegg blir genomiske verktøy faset inn for å øke effektiviteten i skogplanteforedlingen. Alle disse brikkene kan bidra til å snevre inn den genetiske diversiteten, men først og fremst anrike gunstige gener for de egenskapene som vektlegges i foredlingen
– tilvekst, klimatilpasning og kvalitet.
VARIASJON MÅ VÆRE SOM I NATUREN
Genetisk diversitet i foredlet gran er ikke noe vi kan ta lett på. Det er stor forskjell på å foredle treslag som finnes naturlig i store populasjoner og skal tjene en rekke forskjellige formål, og hvete som en innført og domestisert art med ett formål
– å bli mat for folk og fe. Dette er bakgrunnen for synet om at den genetiske diversiteten i foredlet materiale av gran må ligge innenfor variasjonsområdet for naturlige populasjoner, også i fremtiden. Gode referansematerialer og moderne genetiske metoder gjør det mulig å overvåke genetisk diversitet over tid. Vi er også avhengig av å hele tiden supplere med ny forskning som gir et skarpere bilde av foredlingens genetiske effekter, for eksempel på tilpasningsegenskaper. Dersom det skulle være behov for å supplere med genetisk variasjon kan dette gjøres fra arkivmaterialer som Skogfrø verket bevarer.